
Introduction to the Mesenchymal-Epithelial Transition (MET)
A mesenchymal-to-epithelial transition (MET) and its reverse process epithelial-to-mesenchymal transition (EMT) are evolutionarily conserved programmes involved in embryogenesis [1,2], wound healing [2,3], and cancer metastasis [2,4]. MET is characterized by a complex phenotypic change from motile, spindle-shaped mesenchymal cells into sheets of polarized epithelial cells [5].
MET involves upregulation of junctional complexes required for cell adhesion to other cells and to the extracellular matrix (ECM) [1]. Intercellular adhesion is established through the formation of adherens junctions, tight junctions, gap junctions, and desmosomes along the lateral membrane [1,6], while interaction with the ECM is primarily mediated through upregulation of integrins existing at the basal membrane [1,6,7] (Fig. 1).
Together, intercellular adhesion and cell-ECM interactions initiate the process of polarization during MET [5]. Reorganization of the actin cytoskeleton coordinates endosomal trafficking, helping to establish apical-basal polarity [1]. Specifically, apical-basal polarity refers to unequal distribution of proteins and intracellular organelles between the apical and basal sides of epithelial cells [5]. For example, cells undergoing MET localize the Golgi apparatus to the apical cytoplasm and situate their basal cell surface on top of the basement membrane [7].
MET ultimately replaces loose mesenchymal cells with sheets of connected epithelial cells that bear a distinct phenotype [1,2,5]. Due to polarization and upregulation of junctional complexes, cells undergoing MET downregulate mesenchymal markers such as neural cadherin (N-cadherin), collagen type I and III, and vimentin [8]. Instead, cells upregulate their expression of epithelial surface markers such as epithelial cadherin (E-cadherin), collagen type IV and VII, tight junction protein 1 (TJP1/ZO-1), and laminin [9].
Metaplasia and MET
The term ‘metaplasia’ was originally created based on anatomical observations of foreign tissues existing at ectopic sites [10]. Today, metaplasia is defined as the replacement of one differentiated cell type with another differentiated cell type not normally expressed in a specific tissue [11]. A subset of metaplasia known as transdifferentiation (TD) refers to the direct conversion between differentiated cell types without going through a dedifferentiated “intermediate” (i.e., an intermediate pluripotent state or progenitor cell type) [12].
The process of MET, which is crucial to both embryogenesis [2] and the metastasis of secondary tumors [6], bears similarities to TD. Recently, Wang et al. demonstrated that culturing metastatic breast and prostate cancer cells on nanostructures significantly promoted MET [13]. However, these researchers also found that nanostructure-cultured cells acquired expression of both epithelial and mesenchymal markers [13]. Other evidence also shows embryonic and cancer cells displaying attributes of both epithelial and mesenchymal phenotypes, supporting the existence of a transient intermediate existing in the process of MET [14,15,16,17].
Likewise, an intermediate cell phenotype has also been discovered in the process of TD, with researchers finding that TD can only reach completion upon passing through this intermediate state [18]. The possession of an intermediate phenotype in both MET and TD may point to similarity between the two processes.
However, TD and metaplasia both replace a differentiated cell type with another cell type derived from the same embryonic germ layer [12,19]. In contrast, MET replaces cells with cell types derived from a different germ layer [1,2,3,6]. Mesenchymal cells, derived from the mesoderm, undergo MET to be replaced by epithelial cells, which are instead derived from the ectoderm/endoderm — entirely different germ layers [20].
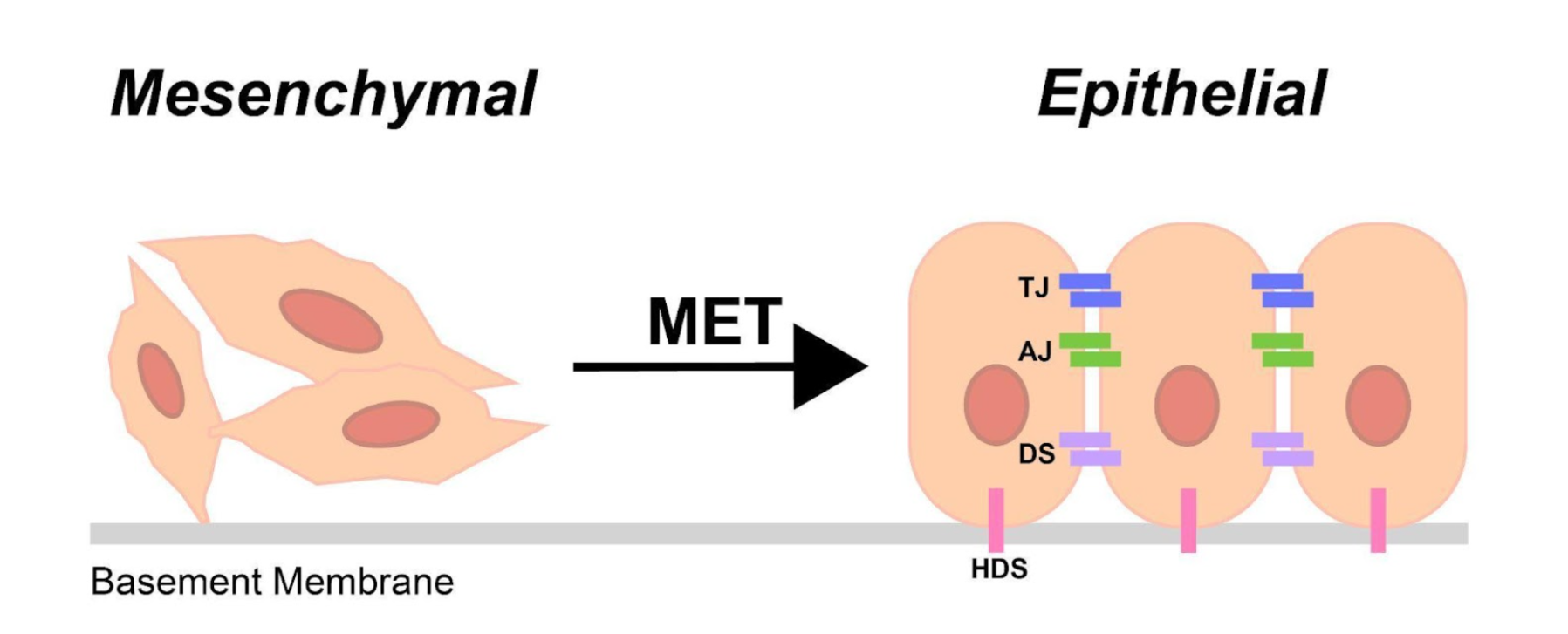
Figure 1: The mesenchymal-epithelial transition (MET): Mesenchymal cells undergoing MET upregulate epithelial junctional complexes required for cell-cell and cell-ECM adhesion. Tight junctions (TJ), adherens junctions (AJ), gap junctions (GJ), and desmosomes (DS) are formed along the lateral membrane, and hemidesmosomes (HDS) are formed along the basal membrane.
Additionally, MET often replaces undifferentiated progenitors as opposed to terminally differentiated cell types [9,21,22]. MET does not usually replace one differentiated cell type with another differentiated cell type1, [9,21,22], making it difficult to explicitly define the process as a form of metaplasia. Rather, MET often involves the reprogramming of undifferentiated cells such as mesenchymal stem cells (MSCs) [21], embryonic stem cells (ESCs) [1], and cancer stem cells (CSCs) [2,22].
Numerous examples of MET occurring across embryonic germ layers have been observed. For example, bone marrow-derived mesenchymal stem cells (BM-MSCs) are adult stem cells that have been shown to populate endo/ectoderm-derived tissues and convert into epithelial cells of the liver, kidney, lung, and pancreas through MET [23,24,25,26]. Engraftment of BM-MSCs into hepatocytes (epithelial cells) following bone marrow transplantation was first observed in response to liver damage [24]. To test whether liver damage promoted the MET of MSCs into hepatocytes, researchers administered hepatotoxin (induces liver damage) and 2-acetylaminofluorine (prevents endogenous liver repair) to rats [24,27]. Using fluorescence in situ hybridization (FISH), resultant hepatocytes were confirmed to be derived from the transplanted BM-MSCs, suggesting that the developmental potential of BM-MSCs is not restricted to their germ layer of origin [24]. Even during nephrogenesis, cells of the mesenteric mesenchyme undergo MET to form the renal epithelium [2,28,29]. Likewise, carcinoma cells often undergo MET to efficiently metastasize at distant tissue sites, changing from mesenchymal to epithelial cells [30]. In each of these examples, MET is shown to occur across germ layers or involve stem cells, features that do not comply with consensus definition of metaplasia [11].
Moving Forward with Understanding MET
The mechanism of MET is poorly characterized compared to EMT [1,2,3,4,5], and several unknowns still exist in the field of epithelial and mesenchymal transitions. It is poorly understood how epithelial cells derived from different types of mesenchymal cells differ on the molecular level. MSCs and fibroblasts (FB) represent two distinct types of mesenchymal cells that are both known to undergo MET [9,21]. An experiment that could help elucidate the differences between MSC-derived and fibroblast-derived epithelial cells could be done by culturing MSCs and FB separately on nanostructures known to promote MET [13]. After MET is induced by the nanostructures, cells could be incubated with an antibody that binds to E-cadherin, a marker specific to epithelial cells [9]. Then, antibody-bound cells could be isolated using fluorescence-activated cell sorting (FACS). Genetic and transcriptomic analyses such as next-generation sequencing (NGS) and single-cell RNA sequencing (RNA-seq) could be conducted on these isolated epithelial cells to observe molecular differences existing between epithelial cells derived from either MSCs or fibroblasts. However, substantial research will be required to properly translate current knowledge on MET to a clinical setting.
Acknowledgements
Declaration of Competing Interests
The author has no competing interests to declare.
Author Information
Author and Affiliations
University of Toronto, Toronto, ON, Canada
Samuel Kogan
Corresponding Author
Please direct correspondence to Samuel Kogan.
References
- 1. Pei, D., Shu, X., Gassama-Diagne A., & Thiery, J. Mesenchymal-epithelial transition in development and reprogramming. Nat Cell Biol. 21, 44-53 (2019).
- 2. Yang, J., Antin, P., Berx, G., Blanpain, C., Brabletz, T., Bronner, M., Campbell, K., Cano, A., Casanova, J., Christofori, G., Dedhar, S., Derynck, R., Ford, H. L., Fuxe, J., García de Herreros, A., Goodall, G. J., Hadjantonakis, A. K., Huang, R. J. Y., Kalcheim, C., Kalluri, R., Kang, Y., Khew-Goodall, Y., Levine, H., Liu, J., Longmore, G. D., Mani, S. A., Massagué, J., Mayor, R., McClay, D., Mostov, K. E., Newgreen, D. F., Nieto, M. A., Puisieux, A., Runyan, R., Savagner, P., Stanger, B., Stemmler, M. P., Takahashi, Y., Takeichi, M., Theveneau, E., Thiery, J. P., Thompson, E. W., Weinberg, R. A., Williams, E. D., Xing, J., Zhou, B. P., & Sheng, G. Guidelines and definitions for research on epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 21, 341-352 (2020).
- 3. Kalluri, R. EMT: When epithelial cells decide to become mesenchymal-like cells. J. Clin. Investig. 119, 1417-1419 (2009).
- 4. Pastushenko, I., Brisebarre, A., Sifrim, A., Fioramonti, M., Revenco, T., Boumahdi, S., van Keymeulen, A., Brown, D., Moers, V., Lemaire, S., de Clercq, S., Minguijón, E., Balsat, C., Sokolow, Y., Dubois, C., de Cock, F., Scozzaro, S., Sopena, F., Lanas, A., D’Haene, N., Salmon, I., Marine, J. C., Voet, T., Sotiropoulou, P. A., & Blanpain, C. Identification of the tumour transition states occurring during EMT. Nature 556, 463-468 (2018).
- 5. Lamouille, S., Xu, J., & Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 15, 178-196 (2014).
- 6. Dongre, A., Weinberg, R.A. New insights into the mechanisms of epithelial–mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 20, 69–84 (2019).
- 7. Hay, E.D. Transformations between epithelium and mesenchyme: normal, pathological, and experimentally induced. Am. J. Kidney Dis. 26, 678-690 (1995).
- 8. Das, V., Bhattacharya, S., Chikkaputtaiah, C., Hazra, S., & Pal, M. The basics of epithelial–mesenchymal transition (EMT): A study from a structure, dynamics, and functional perspective. J. Cell Physiol. 243, 14535-14555 (2019).
- 9. Becerril, C., Montaño, M., Cisneros, J., Mendoza‐milla, C., Pardo, A., Ortiz‐quintero, B., Selman, M., & Ramos, C. Mesenchymal–epithelial transition in fibroblasts of human normal lungs and interstitial lung diseases. Biomolecules 11, 1–21. (2021).
- 10. Slack, J.M.W. Epithelial metaplasia and the second anatomy. Lancet 2, 268-271 (1986).
- 11. Giroux, V., & Rustgi, A. K. Metaplasia: Tissue injury adaptation and a precursor to the dysplasia-cancer sequence. Nat. Rev. Cancer 17, 594-604 (2017).
- 12. Thowfeequ, S., Myatt, E. J., & Tosh, D. Transdifferentiation in developmental biology, disease, and in therapy. Dev. Dyn. 236, 3208–3217 (2007).
- 13. Wang, Z., Xia, F., Labib, M., Ahmadi, M., Chen, H., Das, J., Ahmed, S. U., Angers, S., Sargent, E. H., & Kelley, S.O. Nanostructured Architectures Promote the Mesenchymal-Epithelial Transition for Invasive Cells. ACS Nano 14, 5324–5336 (2020).
- 14. Rodriguez-Boulan, E. & Macara, I. G. Organization and execution of the epithelial polarity programme. Nat. Rev. Mol. Cell Biol. 15, 225–242 (2014).
- 15. Dong, J., Hu, Y., Fan, X., Wu, X., Mao, Y., Hu, B., Guo, H., Wen, L., & Tang, F. Single-cell RNA-seq analysis unveils a prevalent epithelial/mesenchymal hybrid state during mouse organogenesis. Genome Biol. 19, 31 (2018).
- 16. Huang, R. Y., Guilford, P. & Thiery, J. P. Early events in cell adhesion and polarity during epithelial–mesenchymal transition. J. Cell Sci. 125, 4417–4422 (2012).
- 17. Lim, J. & Thiery, J.P. Epithelial-mesenchymal transitions: insights from development. Development 139, 3471–3486 (2012).
- 18. Ye, Z., Li, W., Jiang, Z., Wang, E., & Wang, J. An intermediate state in trans-differentiation with proliferation, metabolic, and epigenetic switching. iScience 24, 103057 (2021).
- 19. Slack, J.M.W. Metaplasia and somatic cell reprogramming. J. Pathol. 217, 161-168 (2009).
- 20. Itskovitz-Eldor, J., Schuldiner, M., Karsenti, D., Eden, A., Yanuka, O., Amit, M., Soreq, H., & Benvenisty, N.. Differentiation of Human Embryonic Stem Cells into Embryoid Bodies Comprising the Three Embryonic Germ Layers. Mol. Med. 6, 88–95 (2000).
- 21. Fujioka, N., Kitabatake, M., Ouji-Sageshima, N., Ibaraki, T., Kumamoto, M., Fujita, Y., Hontsu, S., Yamauchi, M., Yoshikawa, M., Muro, S., & Ito, T. Human adipose-derived mesenchymal stem cells ameliorate elastase-induced emphysema in mice by mesenchymal-epithelial transition. Int. J. Chronic Obstr. Pulm. Dis. 16, 2783–2793 (2021).
- 22. Majumdar, S., & Liu, S. T. Cell division symmetry control and cancer stem cells. AIMS Mol. Sci. 7, 82–98 (2020).
- 23. Heike, T., & Nakahata, T.. Stem cell plasticity in the hematopoietic system. Int. J. Hematol. 79, 7-14 (2004).
- 24. Herzog, E. L., Chai, L., & Krause, D. S. Plasticity of marrow-derived stem cells. Blood 102, 3483–3493 (2003).
- 25. Alison, M., Poulsom, R., Jeffery, R., Dhillon, A. P., Quaglia, A., Jacob, J., Novelli, M., Prentice, G., Williamson, J., & Wright, N. A. Hepatocytes from non-hepatic adult stem cells. Nature 406, 257 (2000).
- 26. Petersen, B. E., Bowen, W. C., Patrene, K. D., Mars, W. M., Sullivan, A. K., Murase, N. A., Boggs, S. S., Greenberger, J. S., & Goff, J. P. Bone marrow as a potential source of hepatic oval cells. Science 284, 1168-1170 (1999).
- 27. Evarts, R. P., Nagy, P., Nakatsukasa, H., Marsden, E., & Thorgeirsson, S. S. In Vivo Differentiation of Rat Liver Oval Cells into Hepatocytes. Cancer Research 49, 1541-1547 (1989).
- 28. Dressler, G. R. The cellular basis of kidney development. Annu. Rev. Cell Dev. Biol. 22, 509–529 (2006).
- 29. Stark, K., Vainio, S., Vassileva, G., & McMahon, A. P. Epithelial transformation of metanephric mesenchyme in the developing kidney regulated by Wnt-4. Nature 372, 679–683 (1994).
- 30. Pattabiraman, D. R., Bierie, B., Kober, K. I., Thiru, P., Krall, J. A., Zill, C., Reinhardt, F., Tam, W. L., & Weinberg, R. A. Activation of PKA leads to mesenchymal-to-epithelial transition and loss of tumor-initiating ability. Science 351, aad3680 (2016).

